ANATOMY OF THE SKIN
Maged N. Kamel, M.D.
ANATOMY OF THE SKIN
Maged N. Kamel, M.D.
Sensory Apparatus of the Skin
The skin is innervated with around one million afferent nerve
fibers. Most terminate in the face and extremities; relatively few
supply the back. The cutaneous nerves contain axons with cell bodies
in the dorsal root ganglia. Their diameters range from 0.2-20
µm. The main nerve trunks entering the subdermal fatty tissue
each divide into smaller bundles. Groups of myelinated fibers fan out
in a horizontal plane to form a branching network from which fibers
ascend, usually accompanying blood vessels, to form a mesh of
interlacing nerves in the superficial dermis. Throughout their
course, the axons are enveloped in Schwann cells and as they run
peripherally, an increasing number lack myelin sheaths. Most end in
the dermis; some penetrate the basement membrane, but do not travel
far into the epidermis. ![]()
Sensory endings are of two main kinds: corpuscular, which embrace
non-nervous elements, and 'free', which do not. Corpuscular endings
can, in turn, be subdivided into encapsulated receptors, of which a
range occurs in the dermis, and non-encapsulated, exemplified by
Merkel's 'touch spot' which is epidermal. ![]()
Each Merkel's touch spot is composed of a battery of Merkel cells
borne on branches of a myelinated axon. A Merkel cell has a lobulated
nucleus and characteristic granules; it is embedded in the basal
layer of epidermal cells, with which it has desmosomal connections;
it contains intermediate filaments composed of low molecular weight
keratin rather than neurofilament protein. ![]()
The Pacinian corpuscle is one of the encapsulated receptors. It is an
ovoid structure about 1mm in length, which is lamellated in
cross-section like an onion, and is innervated by a myelinated
sensory axon which loses its sheath as it traverses the core. The
Golgi-Mazzoni corpuscle found in the subcutaneous tissue of the human
finger is similarly laminate but of much simpler organization. These
last two lamellated end organs are movement and vibration
detectors. ![]()
The Krause end bulb is an encapsulated swelling on myelinated fibers
situated in the superficial layers of the dermis. Meissner corpuscles
are characteristics of the papillary ridges of glabrous (hairless
skin) skin; they are touch receptors; they have a thick lamellated
capsule, 20-40 µm in diameter and up to 150 µm long.
Ruffini endings in the human digits have several expanded endings
branching from a single myelinated afferent fibre; the endings are
directly related to collagen fibrils; they are stretch
receptors. ![]()
'Free nerve endings', which appear to be derived from non-myelinated
fibers occur in the superficial dermis and in the overlying
epidermis; they are receptors for pain, touch, pressure and
temperature. Hair follicles have fine nerve filaments running
parallel to and encircling the follicles; each group of axons is
surrounded by Schwann cells; they mediate touch sensation.
![]()
The brain receives two types of sensations: (1) superficial
sensations, including pain, temperature and crude touch, and (2) deep
sensations, including sense of position, sense of movement, vibration
sense, muscle sense and fine touch. Some superficial and deep
sensations must reach the cortex to be felt. These are: (1) tactile
localization, tactile discrimination and stereognosis, (2) mid-zones
of temperarure (between very hot and very cold values), and (3) sense
of position and movement. ![]()
Pathway of pain, temperature and crude touch sensations: (1)
The first order neuron is present in the posterior root ganglion. Its
dendrite passes to the periphery to act as a pain receptor, while its
axon passes towards the spinal cord. In the spinal cord, it ascends
for one or few segments at the tip of the posterior horn forming the
Lissauer's tract, and then ends around the cells of the substantia
gelatinosa of Rolandi which are present at the tip of the posterior
horn of the grey matter. (2) The second order neuron is present in
the substantia gelatinosa of Rolandi. Its axon crosses to the
opposite side in the anterior commissure near the central canal, then
ascends in the lateral spinothalamic tract (ventral spinothalamic
tract, in case of crude touch) to terminate in the thalamus. (3) The
third order neuron is present in the thalamus. Its axon travels in
the posterior limb of the internal capsule behind the pyramidal
fibers and terminates in the sensory area of the cerebral cortex
(areas 1, 2, and 3). ![]()
Pathway of deep sensations and fine touch: (1) The first order
neuron is also present in the posterior root ganglion. Its dendrite
passes to the periphery, while its axon enters the spinal cord and
ascends directly (without relay) in the posterior column of the
spinal cord, forming the Gracile and Cuneate tracts. The two tracts
end in the medulla around the Gracile and Cuneate nuclei. (2) The
second order neuron is present in the Gracil and Cuneate nuclei of
the medulla. Its axon crosses to the opposite side then ascends in
the brain stem (forming the medial lemniscus) to reach the thalamus
where it terminates. (3) The third order neuron is present in the
thalamus. Its axon passes upwards in the internal capsule to end in
the sensory area of the cerebral cortex. ![]()
Physiology of Sensory Receptors
Adaptation: When a maintained stimulus of constant strength
is applied to a receptor, the frequency of the action potentials in
its sensory nerve declines over time. There are two types of
receptors: (1) tonic slowly-adapting receptors: as the nociceptors
(pain receptors) which continue to transmit impulses to the brain as
long as the stimulus is applied, thus keeping the CNS continuously
informed about the state of the body; and (2) phasic rapidly-adapting
receptors: as Pacinian corpuscles--these receptors adapt rapidly and
cannot be used to transmit a continuous signal to the CNS -they are
stimulated only when the stimulus strength is changed.
![]()
Touch sensation is provoked by a harmless stimulus to the skin
allowing us to distinguish between hard and soft objects; touch
receptors belong to the class of mechanoreceptors and many of them
can be found around hair follicles, so removal of hair decreases
touch sensitivity; the tips of the fingers and lips are rich in touch
receptors. ![]()
Tickle and itch: These sensations are experienced when mild
stimulation of the pain nerve endings occurs; there are also specific
free nerve endings for itch sensation; tickle and itch sensations are
transmitted by group C unmyelinated nerve fibers; histamine produces
itch while pain signals suppress it; tickle is itch produced by light
external moving stimuli and is a pleasurable sensation; itch is an
annoying sensation while pain is unpleasant; itch sensation excites
the scratch reflex. ![]()
Endorphins and enkephalins are important opioid neurotransmitters in
the CNS that mediate the sensation of itch. Although morphine
alleviates pain, it aggravates itch, as itch and pain share common
neurological pathways. The central elicitation of itch by morphine
results from binding to opioid receptors and this binding may mimic
normal physiological binding of endorphins and enkephakins at these
receptor sites. Naloxone, an opioid antagonist, has been found to
reduce histamine-provoked itch. ![]()
The heparin-containing tissue cells called mast cells have a high
histamine content in their granules. They also contain serotonin.
Mast cells are particularly numerous in the skin (about 7,000 mast
cells per cubic millimeter in normal skin in the subpapillary
dermis). Mast cells play an important role in type I immediate
hypersensitivity reation (IgE-mediated anaphylactic
reaction). ![]() n
,
n
,
Temperature sensation: Receptors for warmth and cold are
specialized free nerve endings; a rise in skin temperature above body
temperature causes a sensation of warmth, while a fall in skin
temperature below body temperature is experienced as cold sensation;
pain is felt if skin temperature increases above 45 °C or
decreases below 10 °C; the mucous membrane of the mouth is less
sensitive than the skin, thus tea can be drunk at a temperature which
is painful to fingers. ![]()
Paradoxical cold: Cold receptors are stimulated by intrinsic heat
(e.g., shivering that occurs with fever). ![]()
Pain is evoked by non-specific stimuli (chemical, mechanical,
thermal, or electrical) of an intensity which can produce tissue
damage. Pain is a high threshold sensation. The nociceptors (pain
receptors) are free nerve endings. Cutaneous pain may be sharp and
localized, or dull and diffuse. A painful stimulus causes at first
sharp pain, followed by dull aching pain. Reflex withdrawal movements
also occur, with an increase in heart rate and blood pressure. Fast
sharp (pricking) pain is mediated by nociceptors innervated by group
A delta thick myelinated nerve fibers which transmit pain impulses at
a velocity of 20 meter/second. Slow chronic (dull-aching or burning)
pain is mediated by nociceptors innervated by group C thin
unmyelinated nerve fibers that conduct pain at a low velocity of 1
meter/second. ![]()
Algogenic substances: These are pain-producing substances either
exogenous (as acids and alkalies), or endogenous (as prostaglandins,
bradykinin, ATP, 5-HT [serotonin], and histamine).
![]()
Types of Hyperalgesia: (1) Primary hyperalgesia: Hypersensitivity of
pain receptors (lowered pain threshold, so that touch can produce
pain). It is due to release of algogenic substances (as histamine and
prostaglandins) in an inflamed area of the skin (e.g., sunburn). (2)
Secondary hyperalgesia: It is not due to a skin lesion. The cause is
a CNS lesion (e.g., thalamic syndrome [see: Cutaneous Sensory
System: Connection to CNS] and herpes zoster) with facilitation
of sensory transmission. The pain threshold in secondary hyperalgesia
is normal or elevated, but the pain produced is unpleasant, prolonged
and severe. ![]()
Circulation through the skin serves two functions: (1) nutrition
of the skin tissue, and (2) regulation of body temperature by
conducting heat from the internal structures of the body to the skin,
where it is lost by exchange with the external environment (by
convection, conduction and radiation). (See Also: Sweat
Glands.) ![]()
The cutaneous circulatory apparatus is well-suited to its functions.
It comprises two types of vessels: (1) the usual nutritive vessels
(arteries, capillaries and veins), and (2) vascular structures
concerned with heat regulation. The latter include an extensive
subcutaneous venous plexus which can hold large quantities of blood
(to heat the surface of the skin), and arteriovenous anastomoses
which are large direct vascular communications between arterial and
venous plexuses. Arteriovenous anastomoses are only present in some
skin areas which are often exposed to maximal cooling, as the volar
surfaces of hands and feet, the lips, the nose and the ear.
![]()
The specialized vascular structures just mentioned, bear strong
muscular coats innervated by sympathetic adrenergic vasoconstrictor
nerve fibers. When constricted, blood flow into the subcutaneous
venous plexus is reduced to almost nothing (minimal heat loss);
while, when dilated, extremely rapid flow of warm blood into the
venous plexus is allowed (maximal heat loss).
![]()
The blood flow required for the nutrition of the skin is very small
(about 40ml/min). Yet, at ordinary skin temperature, the amount of
blood flowing through the skin is 10 times (=0.25L/m2 =400ml/min in a
normal adult) more than what is needed for the nutrition of the
tissues. ![]()
The rate of cutaneous blood flow required to regulate body
temperature varies in response to changes in the metabolic activity
of the body and/or the temperature of the surroundings. Exposure to
extreme cold reduces the rate of blood flow to very low values, so
that the nutritive function may sometimes suffer. On the other hand,
heating the skin until maximal vasodilatation occurs (as in hot
climate), increases the cutaneous blood flow 7 times the normal value
(2.8L/min.). This diminishes the peripheral resistance and increases
the cardiac output, which may lead to the decompensation of the heart
in borderline-heart-failure subjects exposed to hot weather.
![]()
Located in the anterior hypothalamus is a small center that controls
body temperature. Heating this area causes vasodilatation of all the
skin vessels of the body and sweating. On the contrary, cooling this
center causes vasoconstriction of skin vessels and stoppage of sweat
secretion. The hypothalamus exerts its controlling effect on the skin
vessels through sympathetic nerves. There are also vasoconstrictor
reflex centers in the spinal cord. ![]()
Sympathetic noradrenergic vasoconstrictor fibers supply the vessels
of the skin throughout the body. This constrictor system is extremely
powerful in the feet, hands, lips, nose and ears (areas where large
numbers of arteriovenous anastomoses are found). At normal body
temperature, the sympathetic vasoconstrictor nerves keep these
anastomoses closed. However, when the body becomes overheated, the
sympathetic discharge is greatly reduced so that the anastomoses
dilate allowing large quantities of warm blood to flow into the
subcutaneous venous plexus, thereby promoting heat loss from the
body. ![]()
Active vasodilatation of the blood vessels of the skin of forearms
and trunk may be due to the discharge of sympathetic cholinergic
vasodilator fibers supplying these areas. The increased activity of
sweat glands in hot weather may also contribute to the vasodilatation
by releasing kallikrein, an enzyme which splits the polypeptide
bradykinin from a globulin present in the interstitial spaces.
Bradykinin is a powerful vasodilator. ![]()
In cold weather, when the temperature reaches 15 °C, we get
maximal vasoconstriction of skin blood vessels. However, in a normal
subject, if the skin temperature is lowered below 15 °C, the
cutaneous vessels begin to dilate. This dilatation is attributed to
the direct local effect of cold causing paralysis of the contractile
musculosa of skin blood vessels and blocking nerve impulses to the
vessels. Maximal vasodilatation occurs at 0 °C, increasing the
blood flow through the skin which prevents freezing of the exposed
areas of the body. ![]()
The cutaneous circulation also serves as a blood reservoir. Under
conditions of circulatory stress, e.g., exercise and hemorrhage,
sympathetic stimulation of subcutaneous venous plexus forces a large
volume of blood (5-10% of the blood volume) into the general
circulation. ![]()
Reactive hyperaemia occurs if one, for example, sits on one portion
of his skin for 30 minutes or more then removes the pressure. In such
conditions, the individual will notice intense redness of the skin at
the site of previous pressure, which resulted from accumulation of
vasodilator metabolites at that site (due to decreased availability
of nutrients to the tissues during compression).
![]()
The triple response: A firm stroke applied to the skin results
in three local reactions collectively known as the triple response:
(a) at first blanching of the skin occurs for a very brief moment
(due to pressure exerted by the stroke), followed by a red line due
to capillary dilatation (caused by histamine and other mediators of
vasodilatation released from damaged tissues); (b) a red flare
follows the red line by 20-40 seconds and is due to arteriolar
dilatation through a local axon reflex (the axon reflex is caused by
stimulation of the pain nerve fibers with impulses passing up these
fibers and down to their endings where vasodilator algogenic--i.e,
pain-inducing--substances are released); (c) finally, a wheal may
appear after 1 minute, reaching full development within 5 minutes
(the wheal is best seen in people with hypersensitive skin; it is due
to release of histamine which causes arteriolar dilatation and
venular constriction, raising the capillary blood pressure with
transudation of fluid into the tissues). ![]()
The epidermis is a multilayered structure (stratified epithelium)
which renews itself continuously by cell division in its deepest
layer, the basal layer. The principal cell type, the epidermal cell,
is most commonly referred to as a keratinocyte. The cells produced by
cell division in the basal layer constitute the prickle cell layer
and as they ascend towards the surface they undergo a process known
as keratinization which involves the synthesis of the fibrous protein
keratin. The total epidermal renewal time is 52-75 days. The cells on
the surface of the skin, forming the horny layer (stratum corneum),
are fully keratinized dead cells which are gradually abraded by day
to day wear and tear from the environment. ![]()
The basal layer is composed of columnar cells which are anchored to a
basement membrane--this lies between the epidermis and dermis. The
basement membrane is a multilayered structure from which anchoring
fibrils extend into the superficial dermis. Interspersed amongst the
basal cells are melanocytes, large dendritic cells responsible for
melanin pigment production. ![]()
The prickle cell layer acquires its name from the spiky appearance
produced by intercellular bridges (desmosomes) which connect adjacent
cells. Scattered throughout the prickle cell layer are numbers of
dendritic cells called Langerhans cells. Like macrophages, Langerhans
cells originate in the bone marrow and have an antigen-presenting
capacity.
Above the prickle cell layer is the granular layer which is composed
of rather flattened cells containing numerous darkly-staining
particles known as keratohyaline granules. Also present in the
cytoplasm of cells in the granular layer are organelles known as
lamellar granules (Odland bodies). Lamellar granules contain lipids
and enzymes, and they discharge their contents into the intercellular
spaces between the cells of the granular layer and stratum corneum,
providing something akin to 'mortar' between the cellular 'bricks.'
In the granular layer the cell membranes become thickened as a result
of deposition of dense material on their inner surfaces.
![]()
The cells of the stratum corneum are flattened keratinized cells
which are devoid of nuclei and cytoplasmic organelles. These cellular
components degenerate in the upper granular layer. Adjacent cells
overlap at their margins and this locking together of cells, together
with intercellular lipid, forms a very effective barrier. The stratum
corneum varies in thickness depending on the region of the body,
being thickest over the palms of the hands and soles of the
feet. ![]()
The rate of cell production in the germinative compartment of the
epidermis must be balanced by the rate of cell loss at the surface of
the stratum corneum. The control mechanism of epidermopoiesis
consists of a balance of stimulatory and inhibitory signals. Wound
healing provides a model to examine the changes in growth control
that occur in establishing a new epidermis. Wounding of the skin is
followed by a wave of epidermal mitotic activity, which represents
the effects of diffusible factors spreading from the wound into the
surrounding tissue. These factors include cytokines and growth
factors. There production is not limited to immune cells as they are
produced by keratinocytes in vitro and can be found in physiological
amounts in normal human skin. ![]()
Regulation of Epidermopoiesis: Stimulatory Factors
The growth factors which stimulate the epidermal cells include:
epidermal growth factor (EGF), transforming-growth-factor-alpha
(TGFalpha), interleukins (IL) and other immunological cytokines, and
basic fibroblast growth factor (bFGF). ![]()
EGF binds to specific cell-surface receptors (EGFR, a trans-membrane
glycoprotein receptor) present in the basal layer of the human
epidermis. Following binding of EGF to EGFR, the receptor is
internalized and carries EGF into an intracellular cycle within the
cytoplasm and the nucleus to mediate all its effects. EGF has been
shown to increase the growth and persistence of epidermal
keratinocytes and to promote wound healing in vitro. EGF transcripts
are not found in the epidermis, but in salivary glands and intestinal
tract. ![]()
TGFalpha was the first growth factor known to be produced by
keratinocytes. Its mRNA predominates in the basal compartment of the
epidermis. TGFalpha is related to EGF. It binds to and activates the
EGF receptor. It stimulates keratinocyte growth.
![]()
The normal epidermis also contains large amounts of Interleukin-1.
There are two forms, alpha and beta, and unlike macrophages, the
epidermis largely produces IL-1alpha. IL-1 has been shown to be
mitogenic for keratinocytes (other effects include: fibroblast
proliferation and synthesis of collagenase, stimulation of IL-2
production, stimulation of B-cell function, and fever induction).
IL-1 releases IL-6 from keratinocytes. IL-6 appears to stimulate
growth of keratinocytes and can be detected in epidermal cells.
Keratinocytes also synthesize IL-3, IL-4, IL-8 (neutrophil activating
protein), and granulocyte-macrophage colony stimulating
factor. ![]()
Thus the epidermal keratinocytes can, under activation conditions,
secrete a large number of cytokines, which can modulate lymphocyte
activation and granulocyte function. These factors do not work in
isolation but have complex interactions, and may be synergistic or
antagonistic. The factors controlling synthesis and secretion of
these factors may be important in the pathogenesis of skin disease as
well as epidermal growth control. ![]()
The regulation of the effects of growth factors includes the control
of expression of the specific growth-factor receptors. The epidermal
cell cycle is also controlled by the intracellular concentrations of
the cyclic nucleotides: cAMP and cGMP. These are small molecules
which are formed and broken down intracellularly as a response to
external signals acting on the cell membrane. Cyclic AMP is believed
to be the intracellular agent or '2nd messenger' of those hormones,
i.e. catecholamines and polypeptides, which do not themselves
penetrate the surface of cells. Cyclic AMP inhibits epidermal cell
division while cGMP stimulates it. Epidermal mitosis exhibits a
circadian rhythm, which is inversely related to blood adrenaline
levels. ![]()
Steroid hormones like testosterone enter the target cells. Epidermal
keratinocytes contain 5 alpha-reductase enzyme and they can convert
testosterone to 5 alpha-dihydrotestosterone (DHT). DHT binds to
specific cytosol receptors which then translocate to the nucleus,
thereafter, altering protein synthesis via messenger RNA. Androgens
and vitamin A stimulate epidermal mitosis, while glucocorticoid
hormones inhibit it. ![]()
Prostaglandins, which are metabolic products of arachidonic acid, can
affect nucleotide metabolism. Prostaglandins of the D and E series
can increase cAMP, although not all such components are present in
the epidermis. The main prostaglandin formed in the epidermis is
PGE2. On the other hand, lipoxygenase products of arachidonic acid
metabolism namely HETE (12-Hydroxy-Eicosa-Tetra-Enoic acid) and the
leucotrienes are capable of inducing epidermal cell proliferation in
vitro. ![]()
Polyamines, including spermidine, putrescein and spermine, stimulate
mitosis. Ornithine decarboxylase is a particularly important enzyme
for the generation of this group of substances.
![]()
Regulation of Epidermopoiesis: Inhibitory Factors
Growth inhibitors for keratinocytes include chalones,
transforming-growth-factor-beta (TGFbeta), alpha and gamma
interferons (IFN-gamma), and tumour necrosis factor (TNF).
![]()
Chalones are polypeptides produced by suprabasal cells which slow
basal mitosis. TGFbeta stimulates fibroblast growth and increases
fibrosis but inhibits the growth of keratinocytes. Thus although it
may have an inhibitory effect on epidermal growth the effect on wound
healing is complex, because of mesenchymal effects (on fibroblasts)
and it has been reported to stimulate wound healing.
![]()
Alpha and gamma interferons have cytostatic effects on keratinocytes
both in vivo following systemic administration and in vitro.
Following stimulation with IFN-gamma, keratinocytes express class II
antigens, predominantly HLA-DR. High doses of Interferon-gamma are
cytotoxic. ![]()
Thirty percent of administered TNF localizes in epidermis suggesting
the presence of many TNF-binding sites. Keratinocytes also secrete
TNF. TNF can cause release of IL-1. It stimulates fibroblast
proliferation and cytokine production. TNF has also been shown to be
reversibly cytostatic to keratinocytes. ![]()
The skin is structured to prevent loss of essential body fluids,
and to protect the body against the entry of toxic environmental
chemicals. In the absence of a stratum corneum we would all lose
significant amounts of water to the environment, and rapidly become
dehydrated. The stratum corneum with its overlapping cells and
intercellular lipid, makes diffusion of water into the environment
very difficult. ![]()
The skin is also part of the innate immunity (natural resistance) of
the body against invasion by micro-organisms. The dryness and
constant desquamation of the skin, the normal flora of the skin, the
fatty acids of sebum and lactic acid of sweat, all represent natural
defense mechanisms against invasion by micro-organisms. Langerhans
cells present in the epidermis have an antigen-presenting capacity
and might play an important role in delayed hypersensitivity
reactions. They also play a role in immunosurveillance against viral
infections. Langerhans cells interact with neighboring keratinocytes,
which secrete a number of immunoregulating cytokines, and
epidermotropic T-cells forming the skin immune system: SALT (skin
associated lymphoid tissue). ![]()
Melanin pigment of the skin protects the nuclear structures against
damage from ultraviolet radiation. ![]()
The skin is also a huge sensory receptor for heat, cold, pain, touch,
and tickle. Parts of the skin are considered as erogenous zones. The
skin has great psychological importance at all ages. It is an organ
of emotional expression and a site for the discharge of anxiety.
Caressing favors emotional development, learning and growth of
newborn infants. ![]()
The skin is a vital part of the body's temperature regulation system,
protecting us against hypothermia and hyperthermia, both of them may
be fatal (specialized vascular structures of the dermis/insulation by
fat in subcutaneous tissue/evaporation of sweat).
![]()
The skin plays an important role in calcium homeostasis by
contributing to the body's supply of vitamin D. Vitamin D3
(cholecalciferol) is produced in the skin by the action of
ultraviolet light on dehydrocholesterol. It is then hydroxylated in
the liver and kidneys (needs parathyroid hormone to activate
alpha-hydroxylase) to 1,25 dihydroxycholecalciferol, the active form
of vitamin D. This anti-rachitic vitamin acts on the intestine
increasing calcium absorption (through stimulation of synthesis of
calcium-binding proteins in the mucosal cells of the intestine), as
well as on the kidneys promoting calcium reabsorption.
![]()
Fingerprints, the characteristic elevated ridge patterns on the
finger tips of humans, are unique to each individual. The fingers and
toes, the palms of the hands and soles of the feet, are covered with
a system of ridges which form certain patterns. The term
dermatoglyphics is applied to both the configurations of the ridges,
and also to the study of fingerprints. The medicolegal importance of
the ridge patterns of fingerprints, characteristic dermatoglyphic
abnormalities frequently accompany many chromosomal
aberrations. ![]()
Hair performs no vital function in humans, whose body could be
perpetually depilated without any physiological disadvantages. At the
same time the psychological functions are inestimable: scalp hair is
a major social and sexual display feature of the human body.
![]()
Hairs grow out of tubular invaginations of the epidermis known as
follicles, and a hair follicle and its associated sebaceous glands
are referred to as a pilosebaceous unit. Hair follicles extend into
the dermis at an angle. A small bundle of smooth muscle fibers, the
arrector pili muscle, extends from just beneath the epidermis and is
attached to the side of the follicle at an angle. Arrector pili
muscles are supplied by adrenergic nerves, and are responsible for
the erection of hair during cold or emotional stress ('goose flesh').
The sebaceous gland is attached to the follicle just above the point
of attachment of the arrector pili. ![]()
At the lower end of the follicle is the hair bulb, part of which, the
hair matrix, is a zone of rapidly dividing cells which is responsible
for the formation of the hair shaft. Hair pigment is produced by
melanocytes in the hair bulb. Cells produced in the hair bulb become
densely packed, elongated and arranged parallel to the long axis of
the hair shaft. They gradually become keratinized as they ascend in
the hair follicle. ![]()
|
|
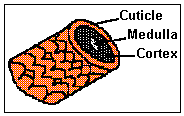
The main part of each hair fibre is the cortex, which is composed of keratinized spindle-shaped cells. Terminal hair (as that of scalp) have a central core known as the medulla consisting of specialized cells which contain air spaces (see: Types of Hair). Covering the cortex is the cuticle, a thin layer of cells which overlap like the tiles on a roof, with the free margins of the cells pointing towards the tip of the hair. |
The cross-sectional shape of hair varies with body site and with
race. African hair is distinctly oval in cross-section, and pubic,
beard and eyelash hairs are oval in all racial types. The form of
scalp hair also differs among human races (e.g., the peppercorn
pattern in black Africans). ![]()
The average rate of growth of human scalp hair is 0.37mm per day. In
women scalp hair grows faster and body hair grows more slowly than in
men. The rate of growth of body hair is undoubtedly increased by
androgens, since it can be reduced by treatment with antiandrogenic
steroids. ![]()
Hair: Types and Growth Cycle
The first hair to be produced by the fetal follicles, so called
lanugo, is fine, soft, unmedullated, and usually unpigmented. Lanugo
is normally shed in utero in the seventh or eighth month of
gestation. ![]()
Postnatal hair can be divided into vellus, which is soft
unmedullated, usually unpigmented, and seldom more than 2cm long, and
terminal hair, which is longer, coarser, and often medullated and
pigmented. There is, however, a range of intermediate types.
![]()
The type of hair produced by any particular follicle can change. The
most striking example is the replacement of vellus by terminal hair
at puberty which starts in the pubic regions. This leads us to the
definition of androgen-dependent hair. It is obvious from the events
of puberty that pubic, axillary, facial, and body hair are
hormone-dependent. So, paradoxically, is pattern baldness (male), in
which terminal hair is replaced by fine, short hair resembling
vellus. The growth of male beard depends on testicular hormones. The
action of testosterone in general involves its reduction to 5
alpha-dihydrotestosterone and binding to an intracellular
receptor. ![]()
The most important feature of hair follicles is that their activity
is intermittent (cyclical). As the hair reaches a definitive length,
it is shed to be replaced by a new hair. Thus a hair follicle will
pass into three stages: an active (anagen) stage, a resting (catagen)
stage, and a telogen stage where the hair stops growing to be finally
shed. In human scalp hair, the anagen stage takes about three years,
the catagen stage takes three weeks, and the telogen phase takes
three months. The hair cycle occurs in different hair follicles
asynchronously, i.e., at a given time, each individual hair follicle
is at a different stage of the hair cycle. ![]()
Electron microscopical examination of cells from all tissues
reveals that they contain a complex, heterogenous, intracytoplasmic
system of filaments. The components of this system include actin,
myosin, and tubulin, whose diameters average approximately 60A°,
150A°, and 250A°, respectively. In addition, other
intracytoplasmic filaments were noted, and since the diameter of
these latter structures was found to be between 70 and 100A°,
they were called intermediate filaments. ![]()
Intermediate filaments form a major part of the cytoskeleton of most
cells and fulfill a variety of roles related to cell shape, spatial
organization, and perhaps informational transfer. The nucleus
contains structures related to these intermediate filaments and many
intracellular components including polyribosomes, mitochondria,
nucleic acids, enzymes, and cyclic nucleotides are attached to the
cytoskeleton. ![]()
Based on their biochemical, biophysical, and antigenic properties, a
number of classes of intermediate filaments can be recognized in
different cell types: desmin (skeletin) in muscle cells, glial
fibrillary acidic filaments in glial cells, neurofilaments in
neurons, vimentin in mesenchymal cells, and keratin in epithelial
cells. In cultured epidermal cells, keratins account for up to 30% of
the cellular protein, while in stratum corneum, keratin accounts for
up to 85% of the cellular protein. ![]()
At least 19 keratin proteins can be identified ranging in molecular
weight from approximately 40,000 to 68,000 micrograms. Moll and his
coworkers published their human keratin catalogue in 1982. According
to this catalogue, there are two keratin subfamilies. The molecular
weight of the members of one (the basic subfamily) is relatively
larger than that of the members of the other (the acidic subfamily).
Each of the keratins is the product of a unique gene and, in
essentially all situations, the keratins are expressed as pairs
containing one member of each subfamily. The two members of each pair
are in the same size rank order within their respective family, e.g.,
the largest acidic keratin is expressed with the largest
basic. ![]()
The type of keratin differs in different tissues, i.e, there are
different types of keratin for keratinized epidermis,
hyperproliferative epidermis of palms and soles, corneal epithelium,
stratified epithelium of the esophagus and cervix, and simple
epithelium of the epidermal glands. As mentioned before, keratin is
the main structural protein of the epidermis.
![]()
The keratinocytes in the basal layer and prickle cell layer
synthesize keratin filaments (tonofilaments) which aggregate into
bundles (tonofibrils). Eventually, in the cells of the stratum
corneum, these bundles of keratin filaments form a complex
intracellular network embedded in an amorphous protein matrix. The
matrix is derived from the keratohyaline granules of the granular
layer. Epidermal keratinization results in the production of a
barrier which is relatively impermeable to substances passing in or
out of the body. ![]()
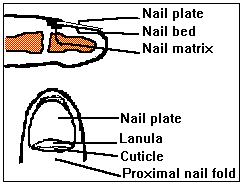
The nail acts as a protective covering to the end of the digit and
assists in grasping small objects. The nail has also a cosmetic
function. The major part of this appendage is the hard nail plate,
which arises from the matrix (see below). The nail plate is roughly
rectangular and flat in shape but shows considerable variation in
different persons. The pink color of the nail bed results from its
extensive vascular network and can be seen because of the
transparency of the plate. ![]()
Usually in the thumbs, uncommonly in other fingers and in the large
toenails, a whitish crescent-shaped lanula is seen projecting from
under the proximal nail folds. The lanula is the most distal portion
of the matrix and determines the shape of the free edge of the nail
plate. Its color is due in part to the effect of light scattered by
the nucleated cells of the matrix and in part to the thick layer of
epithelial cells making up the matrix. ![]()
As the nail plate emerges from the matrix, its lateral and proximal
borders are enveloped by folds of the skin termed the lateral and
proximal nail folds. The skin underlying the free end of the nail is
referred to as the hyponychium and is contiguous with the skin on the
tip of the finger. ![]()
The nail plate is formed by a process which involves flattening of
the basal cells of the matrix, fragmentation of the nuclei, and
condensation of cytoplasm to form horny flat cells which are strongly
adherent to one another. ![]()
|
|
Fingernails grow faster than toenails. Nails of individual fingers
of the same hand grow at different rates. There are also familial
tendencies favouring similar growth rates among persons and it has
been noted that nail growth is increased during summer and diminished
in cold climates. ![]()
Many systemic disorders may produce a decrease in the rate of nail
growth or thinning and grooving of the plate. This phenomenon is best
appreciated weeks after the event has occured. Acute viral infectious
diseases as mumps and measles, starvation, and some types of anaemia
are among the causes. Increase in the growth rate can be seen during
pregnancy, nail biting, trauma and during regrowth after
avulsion. ![]()
Melanocytes and Skin Color
The melanin pigmentary system is composed of functional units
called epidermal melanin units. Each unit consists of a melanocyte
that supplies melanin pigment to a group of keratinocytes (about 36).
Pigmentation is determined primarily by the amount of melanin
transferred to the keratinocytes. ![]()
The melanocyte is a dendritic cell present in the basal layer of the
epidermis with no desmosomes (intercellular bridges) or
tonofilaments. In H & E-stained sections, it has a small dark
nucleus and a clear cytoplasm. It can be stained black with Fontana
Masson (Silver) stain as it contains melanin, and more specifically
with DOPA reaction as it has the ability to form melanin
(tyrosinase-containing cell). ![]()
Melanocytes arise from the neural crest as melanoblasts and migrate
to the dermis, hair follicles, leptomeninges, uveal tract and retina.
By the 8th week of intrauterine life, they start to migrate from the
dermis to the epidermis. Although full melanocyte migration is
normally completed prior to birth, residual dermal melanocytes are
sometimes left (clinically appearing as mongoloid spots in the sacral
area of oriental and black infants). ![]()
Melanosomes are membrane-bound organelles located in the cytoplasm of
melanocytes and bearing tyrosinase enzyme. They are responsible for
melanin synthesis and pigment transfer from the melanocyte to the
surrounding keratinocytes. During their passage from the perinuclear
area of the melanocyte to the dendrites, the melanosomes show four
stages of development: I and II (with no melanin deposition), III
(with high levels of tyrosinase activity and is partially obscured by
melanin deposition), and IV (with low levels of tyrosinase activity
and is completely obscured by melanin deposition). Pigment transfer
occurs by keratinocyte phagocytosis of melanosome-containing
dendritic tips. As squamous cells differentiate, the melanosomes
within them are degraded by lysosomal enzymes.
![]()
The differences in racial pigmentation are not due to differences in
the number of melanocytes, but rather to differences in melanocyte
activity. In black skin, there is greater production of melanosomes,
higher degree of melanization of melanosomes, and larger unaggregated
melanosomes showing slow rate of degradation.
![]()
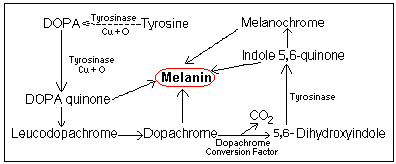
Melanin: Types, Synthesis and Hormonal Regulation
Melanin is a brown-black, light-absorbing pigment, protecting the
skin against ultraviolet rays. Two major forms of melanin exist in
humans: (1) Eumelanin, a brown to black pigment synthesized from
Indole 5,6-quinone and found within the ellipsoid melanosomes; and
(2) Phaeomelanin, a yellow-red pigment found within the spherical
melanosomes. ![]()
|
Tyrosinase is synthesized by the ribosomes of the rough endoplasmic reticulum (rER) and transported through the smooth endoplasmic reticulum (sER) to the Golgi apparatus. It is then released within membrane-bound vesicles. Meanwhile, structural melanosomal proteins are also synthesized on the rER and are then incorporated into vesicles at the sER. Fusion of the two types of vesicles (tyrosinase and structural melanosomal proteins) results in the formation of a melanosome. As the melanosome matures and more melanin is deposited on its lamellar matrix, it passes into the dendrite of the melanocyte. |
|
There are receptors on the surface of melanocytes for
melanocyte-stimulating hormone (MSH). MSH, ACTH (similar to MSH in
the arrangement of first 13 amino acids), estrogen and progesterone,
all stimulate pigmentation through increasing cAMP and tyrosinase
activity, resulting in increased melanin formation and
transfer. ![]()
Sebaceous glands are found on all areas of the skin with the
exception of the palms, soles, and dorsa of the feet. They are
holocrine glands, i.e., their secretion is formed by complete
destruction of the cells. ![]()
Most sebaceous glands have their ducts opening into hair follicles
(pilosebaceous apparatus). Free sebaceous glands (not associated with
hair follicles) open directly to the surface of the skin, e.g.,
Meibomian glands of the eyelids, Tyson's glands of the prepuce, and
free glands in the female genitalia and in the areola of
nipples. ![]()
The production of sebum is under hormonal control and sebaceous
secretion is a continuous process. Sebaceous gland development is an
early event in puberty, and the prime hormonal stimulus for this
glandular development is androgen. Although the sebaceous glands are
very small throughout the prepubertal period, they are large at the
time of birth, probably as a result of androgen stimulation in utero,
and acne may be seen in the neonatal period. It should be noted that:
(1) sebum production is low in children; (2) in adults, sebum
production is higher in men than in women; (3) in men, sebum
production falls only slightly with advancing age, whereas in women
it decreases significantly after the age of 50. Orchidectomy causes a
marked decrease in sebum production. Therefore, it can be assumed
that testicular androgen maintains sebum production at high levels in
men. The role of adrenal androgens is also important, specially in
women where they play a contributory role in sebum production
together with the ovaries. ![]()
Estrogens have a profound effect on sebaceous gland function which is
opposite of that of androgens. In both sexes, estrogen administration
decreases the size of the sebaceous glands and the production of
sebum. ![]()
The sebum is composed of triglycerides and free fatty acids, wax
esters, squalene, and cholesterol. The sebum controls moisture loss
from the epidermis. The water-holding power of cornified epithelium
depends on the presence of lipids. The sebum also protects against
fungal and bacterial infections of the skin due to its contents of
free fatty acids. Ringworm of scalp becomes rare after
puberty. ![]()
Generalized sweating is the normal response to exercise or thermal
stress by which human beings control their body temperature through
evaporative heat loss. Failure of this mechanism can cause
hyperthermia and death. (See Also: Cutaneous Vascular
System.) ![]()
Humans have several million eccrine sweat glands distributed over
nearly the entire body surface (except labia minora and glans penis).
The total mass of eccrine sweat glands roughly equals that of one
kidney, i.e., 100g. A person can perspire as much as several litres
per hour and 10 litres per day, which is far greater than the
secretory rates of other exocrine glands such as the salivary and
lacrimal glands and the pancreas. ![]()
Each eccrine sweat gland consists of a secretory coil deep in the
dermis, and a duct which conveys the secreted sweat to the surface.
The secretory activity of the human eccrine sweat glands consists of
two major functions: (1) secretion of an ultrafiltrate of a
plasmalike precursor fluid by the secretory coil in response to
acetylcholine released from the sympathetic nerve endings, and (2)
reabsorption of sodium in excess of water by the duct, therby
producing a hypotonic skin surface sweat. Under extreme conditions
where the amount of perspiration reaches several litres a day, the
ductal reabsorptive function assumes a vital role in maintaining
homeostasis of the entire body. ![]()
In addition to the secretion of water and electrolytes, the sweat
glands serve as excretory organ for heavy metals, organic compounds,
and macromolecules. The sweat is composed of 99% water, electrolytes,
lactate (provides an acidic pH to resist infection), urea, ammonia,
proteolytic enzymes, and other substances. ![]()
There is a hypothalamic preoptic sweat center that plays an essential
role in regulation of body temperature. Sweat secretion on palms and
soles is more or less continuous (perpetual sweating) when humans are
awake. In contrast, those glands on the general skin surface respond
predominantly to thermal stimuli (thermal sweating). Both types of
sweating can be inhibited by atropine as all sweat glands in
different areas of the body are stimulated by the same sympathetic
cholinergic mechanism. Sweating induced by emotional stress
(emotional sweating) can occur over the whole skin surface, but
usually it is confined to palms, soles, axillae, and the
forehead. ![]()
The term apocrine glands was given to sweat glands present in the
axillae and anogenital area which are under the control of sex
hormones, mainly androgens. But nowadays by electron microscopy,
these apocrine glands (apocrine = apical part of the cell is
destroyed during the process of secretion) proved to be merocrine in
nature (merocrine = no destruction of the cell during the process of
secretion). The "apocrine" sweat of humans has been described as
milky (because it is mixed with sebum due to shared duct) and viscid,
without odour when it is first secreted. Subsequent bacterial action
is necessary for odour production. Unlike eccrine glands which have a
duct that opens onto the skin surface independently of a hair
follicle (atrichial), apocrine glands have a duct that opens into a
hair follicle (epitrichial). ![]()
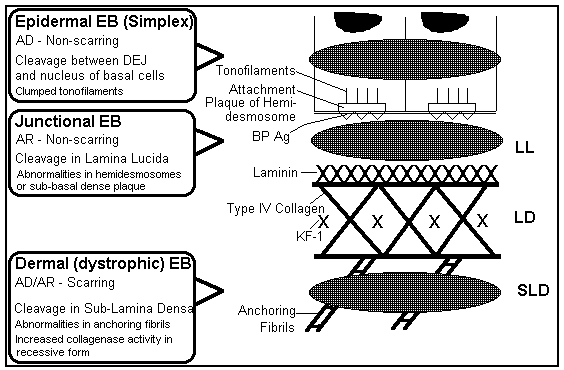
Ultrastructure of the Dermo-Epidermal Junction
The most superficial component of the junction is the basal plasma
membrane of keratinocytes, melanocytes and Merkel cells.
![]()
Hemidesmosomes superficially resemble focal thickening of the basal
plasma membrane of keratinocytes. At higher magnification, however,
they can be seen to have a complicated ultrastructure which resembles
half a desmosome. Hemidesmosomes consist of an intracellular
component, the attachment plaque, which is associated with
tonofilaments, and an extracellular component, known as the sub-basal
dense plate. This latter structure is located in the lamina lucida
(see below) and resembles a fine, dense line parallel to and just
beneath the plasma membrane. Hemidesmosomes are important in
maintaining adhesion between dermis and epidermis.
![]()
Immediately beneath the basal plasma membrane is the basement
membrane which consists of three layers: the lamina lucida, the
lamina densa and the lamina fibroreticularis (sub-lamina
densa). ![]()
Distributed throughout the lamina lucida are anchoring filaments.
Anchoring filaments are very fine structures that are oriented
vertically between the lamina densa and basal plasma
membrane. ![]()
The lamina densa is an electron dense amorphous layer that lies
parallel to and below the lamina lucida. ![]()
Anchoring fibrils are the major constituent of the fibroreticular
layer of the basement membrane. These are short, often curved
structures, with an irregular cross-banding, that insert into the
lamina densa and extend into the upper part of the dermis. They may
also insert into amorphous bodies in the superficial dermis known as
anchoring plaques, or curve back to have a second insertion in the
lamina densa. ![]()
Another component of the lamina fibroreticularis are the elastic
microfibril bundles, each consisting of many microfibrils that extend
into the dermis and may enmesh with the microfibrillar system of
dermal elastic fibers. ![]()
Bullous pemphigoid antigen is a glycoprotein synthesized by
keratinocytes and recognized by circulating autoantibodies in
patients with bullous pemphigoid; it has been localized to
hemidesmosomes--mainly intracellularly, but to a lesser degree, also
just outside the cells. ![]()
Laminin, a high-molecular-weight glycoprotein required for cell
adhesion, has been immunolocalized to lamina lucida. Fibronectin has
also been immunolocalized to the lamina lucida..
![]()
Type IV collagen and KF-1 antigen have been immunolocalized to the
lamina densa. ![]()
Type VII collagen and Epidermolysis bullosa acquisita (EBA) antigen
have been immunolocalized to anchoring fibrils and plaques. Type VII
collagen has a role in normal dermo-epidermal adherence. AF1 and AF2
antigens have also been immunolocalized to the anchoring
fibrils. ![]()
|
|
|
Figure 1: Diagram of the ultrastructure of the dermo-epidermal junction showing the sites of cleavage (blister formation) in the three main types of epidermolysis bullosa, a genetically-determined bullous disorder. |
EB, Epidermolysis Bullosa; AD, Autosomal Dominant; AR, Autosomal
Recessive; DEJ, Dermo-Epidermal Junction; BP Ag, Bullous Pemphigoid
Antigen; LL, Lamina Lucida; LD, Lamina Densa; KF-1, an antigen that
has been immunolocalized to the lamina densa; SLD, Sub-Lamina Densa.
((c) 1994 Dr. Maged N. Kamel) ![]()
Champion RH, Burton JL and Ebling FJG (eds): Textbook of
Dermatology (Rook), 5th ed, 4 vol, Blackwell, Oxford, 1992
Fitzpatrick TB, Eisen AZ, Wolff K, Freedberg IM and Austen KF (eds):
Dermatology in General Medicine, 3d ed, 2 vol, McGraw-Hill, New York,
1987
Ganong WF: Review of Medical Physiology, 14th ed, Appleton &
Lange, Connecticut, 1989
Graham-Brown R and Burns T: Lecture Notes on Dermatology, 6th ed,
Blackwell, Oxford, 1990 ![]()